Биологическое рассеивание
_Taraxacum_sect._Ruderalia.jpg)
Биологическое расселение — постепенное расширение области обитания (ареала) тех или иных видов живых организмов. В результате расселения происходит перемещение животных или рассеивание семян и спор растений, грибов и бактерий с последующим их закреплением (натурализацией) и размножением в новой местности[1][2].
В случае животных расселение связано с изменением условий их проживания и численности животных. Увеличиение численности популяции обуслваливает возрастание плотности проживания особей и усиление внутривидовой конкуренции, что стимулирует расселение животных в новые места обитания[1].
Расселение растений зависит от количества образуемых ими семян и спор (диаспор), выступающих в качестве средств расселения, а также их способностью к закреплению в местах, куда они переносятся (жизнеспособности в новых условиях). При этом выделяют несколько типов расселения в зависимости от движущего фактора, в частности, анемохорию (за счет действия ветра и восходящих потоков воздуха), гидрохорию (за счёт движение водной среды), зоохорию (перенос животными), антропохорию (перенос, связанный с деятельностью человека)[1] и др. типы расселения, изучаемые в рамках диаспорологии — раздела ботаники, занимающегося закономерностями распростронения плодов и семян[3].
На расселение влияет ряд ограничивающих факторов, в частности, географические (реки, моря, проливы, горы), экологические (климатические и другие природные условия), биологические (межвидовая конкуренция). Темп расселения определяется сочетанием действия побуждающих факторов (и средств расселения) и преград, что влияет на динамику численности популяций и геграфическое распределение видов, а также выступает в качестве фактора видовой изменчивости[1][4][5][6][7][8]. Файл:Lichen dispersal.ogv Расселение организмов представляет собой ключевой процесс, влияющий как на эволюционную динамику популяций, так и на пространственное распределение живых организмов. Через перемещение особей и связанный с ним поток генов расселение может ослаблять географическую изоляцию между популяциями либо, напротив, при ограниченном обмене способствовать их дивергенции и видообразованию. Одновременно расселение определяет способность организмов заселять новые территории, расширять или изменять ареалы и поддерживать связь между разобщёнными частями популяций. По этой причине изучение расселения имеет важное значение для понимания общих закономерностей географического распространения организмов, лежащих в основе такого направления биологии как биогеография[9].
Типы расселения[править]
Некоторые организмы имеют возможность к расселению на протяжении всего жизненного цикла, тогда как другие способны к перемещению (или к пассивному переносу) лишь на определённых, ограниченных стадиях развития. Эта стадия обычно называется дисперсионной фазой (или фазой расселения). Во втором случае, стратегии, определяющие полный жизненный цикл организма, нередко формируются под влиянием характера и условий именно этой дисперсионной фазы. При этом различают рассление пассивного и активного типа.

Активное расселение[править]
В этом случае организмы перемещаются в среде обитания самостоятельно, что наиболее характерно для животных, обладающих способностью к свободному перемещению на большие расстояния (бег, плавание, полёт). Возраст при этом не является ограничивающим фактором, поскольку смена местоположения характерна как для молодых, так и для взрослых животных. Активное расселение часто ставят в зависимость от плотности популяции, поскольку именно численность сообщества в привязке к размеру местообитания и качеству среды обитания выступает в качестве движущей силы и определяет масштаб и динамику перемещения животных в новые локации. Плотность популяции может увеличивать нагрузку на ресурсы экосистемы, и расселение способно снижать эту напряжённость; конкуренция за ресурсы, в свою очередь, может выступать фактором отбора, влияющим на формирование механизмов расселения.
Наименьшие ограничения при активном расселении испытывают летающие по воздуху и водоплавающие животные, наибольшие — наземные виды. Хотя некоторые наземные животные способны преодолевать значительные расстояния, передвижение пешком требует больших энергетических затрат по сравнению с полётом или плаванием. Кроме того, их движение может прерываться различными барьерами природного (сложный рельеф, водные преграды) и антропогенного характера (дороги, города)[10]. Обычно различают натальное расселение, при котором особь (как правило, молодая) покидает место своего рождения, и репродуктивное расселение, при котором особь (как правило, взрослая) покидает одно место размножения для размножения в другом.

Пассивное расселение[править]
При пассивном расселении организмы не способны самостоятельно перемещаться, но используют иные механизмы для успешного распространения и заселения новых мест обитания. У таких видов сформировались адаптации к расселению, позволяющие использовать для переноса своих гамет различные формы кинетической энергии, естественным образом возникающие в окружающей среде. Это может происходить за счёт энергии воды, ветра или движения животных, способных к активному перемещению. Некоторые организмы способны к распространению на личиночной стадии развития. Это характерно для ряда беспозвоночных, рыб, насекомых, а также малоподвижных и прикреплённых организмов. Некоторые беспозвоночные, например морские губки и кораллы, высвобождают гаметы в воду. Таким образом осуществляется успешное размножение: сперматозоиды активно перемещаются в воде, тогда как яйцеклетки переносятся течением. Распространение семян и спор, ответственное за расселение растений, происходит за счёт животных-переносчиков, ветра, гравитации или течений. Семена, споры и плоды часто обладают специальными приспособлениями, облегчающими их распространение[11].
Плюсы и минусы расселения[править]
Масштаб и динамика расселения зависят от того, насколько выгоды в ресурсах (пища, территория) и эволюционном развитии, которые получают мигрирующие организмы и популяция в целом, превалируют над негативными последствиями, связанными с временны́ми и энергетическими затратами на перемещение и необходимостью в адаптации на новом месте обитания.
К преимуществам расселения относят освоение новых ресурсов, избегание неблагоприятных условий среды, снижение конкуренции с родственными особями, в том числе сибсами, а также исключение близкородственного скрещивания, следствием которого для изолированных популяций часто является инбредная депрессия[12].
Однако расселение связано с рядом индивидуальных и популяционных рисков, в частности[13]:
- Животным требуется затратить много энергии на перемещение, а растениям — питательных веществ для образования семян и плодов, без гарантии, что мигрирующие особи или семена достигнут нового места с лучшими условиями обитания;
- Для животных, совершающих миграции, существенно повышается риск травм, атак со столроны хищников, риск заражения новыми болезнями;
- В периоды передвижения, животные не могут размножаться и активно расти;
- Переселение может вызвать аутбредную депрессию и обусловленное ею снижение приспособленности потомства (жизнеспособности, фертильности, продуктивности) в результате скрещивания генетически отдаленных особей — например, из разных популяций или подвидов;
- У социальных животных, включая многих птиц и млекопитающих, переселившаяся особь должна суметь интегрироваться в новую популяцию, что может сопровождаться утратой социального статуса.
Факторы расселения: движущие силы и ограничения[править]
Расселение живых существ может сталкиваться с рядом препятствий ландшафтного, антропогенного или экологического характера. Такие барьеры могут приводить к тому, что область фактического расселения вида может оказаться значительно меньше его потенциально возможного ареала или вести к необратимой сегрегации части популяции, которая может выделиться в новый подвид. Факторы расселения могут носить разнонправленный и взаимосвязанный характер, например, антропогенное воздействие может ограничивать расселение животных и выступать в качестве его движущей силы, а также являться причиной появления новых экологических факторов, меняющих картину видового распределения на планете.
Влияние ландшафта: барьеры и векторы расселения[править]
Природный ландшафт[править]

К естественным барьерам, ограничивающим распространение видов, относятся горные хребты, водные объекты (реки, крупные озёра, моря) и другие преграды, связанные с рельефом. Наряду с ограничениями в распространении вида по территории, они могут вызывать разделение видовой популяции и формирование разных особенностей фенотипа и генотипических отличий. Например, обыкновенные шимпанзе и бонобо, это — изначально генетически один вид; но разделённый рекой Конго он эволюционировал в два вида, которые заметно отличаются друг от друга поведением, особенно уровнем агрессии.[14]

Амазонка разделяет ландшафт настолько сильно, что ряд видов обезьян не могут её пересекать.[15] В данный момент в пойме Амазонки сформировались 11 центров, где проживают эндемичные виды, их относительно сильно изолируют 9 рек — Бранко, Негро, Солимойнс, Джавари, Шингу, Мадейра, Токантинс, Тапажос и Амазонка. Появление рек Мадейра и Тапажос способствовало образование новых видов, за счёт изоляции. Трубачи плохо летают, но хорошо бегают и иногда плавают.[16]
Хорошим примером являются птицы-трубачи, получившие свое название из-за угрожающего трубного крика самцов. Эти птицы эндемичны для амазонских лесов, и в настоящее время признано восемь видов. Две группы видов разделились к северу и югу от реки Амазонки 2,7-2,0 миллиона лет назад. Данные свидетельствуют о том, что появление некоторых амазонских рек, таких как Мадейра и Тапажо, изолировало часть птиц по обе стороны реки, что привело к развитию новых видов с течением времени. 30 % из 240 видов птиц, населяющих леса на одном берегу нижнего течения Рио-Негро и Рио Бранко, замещены близкородственными видами на другом берегу. Например на левом берегу Риу Негру есть вид Крапчатый кабезон — Capito niger с красным горлом, а направом берегу живут только близкородственные ему Бородастики-кабезоны — Capito auratus с оранжевым горлом.[17]
В этом же регионе тукан вида гвианская селенидера, разделен рекой с туканом tawny-tufted toucanet (Selenidera nattereri, виды разделились судя по молекулярным часам, около 8 миллионов лет назад.
Городская застройка[править]
Хозяйственная деятельность человека ведёт к изменению природного ландшафта. Это вызывает искусственную фрагментацию территории, потенциально пригодной для расширения видового ареала, за счёт появления городской и сельскохозяйственной инфраструктуры и строений, а также транспортных сетей. Городская застройка вносит свой особый вклад в процессы расселения живых организмов. Для растений городская инфраструктура обеспечивает дополнительные возможности для расселения. В последнее время автомобили стали основным вектором распространения семян растений, наряду с природными способами их распространения (вода, ветер, и т. п.)[18]. Животные могут использовать транспортную инфраструктуру для своего расселения. Например ряд грызунов используют мосты и мелкие переправы через реки, переходя на другой берег, чего ранее делать не могли. Европейские суслики таким образом в 1930-х, смогли расселится из степей Калмыкии на юго-запад[19]. При этом, городсткая среда может вносить и ограничения в отдельные способы расселения. В частности, заметно сужается расстояние полёта насекомых-опылителей, что, в свою очередь, уменьшает обмен генами между растениями популяций, сильно удаленных друг от друга, и, как следствие, снижает их генетическое разнообразие[20]. Ограничения в перемещении животных, например, летучих мышей и мышей, живущих в городах, является фактором, спсобствующим генетической изоляции популяций этих видов[21].
Конкуренция[править]
Наличие или отсутствие видов, занимающих сходные пищевые ниши, — важный фактор, влияющий на успешность расселения вида. Если новый вид попадает в новую среду, но его пищевая ниша совпадает с нишей уже обитающего здесь вида, между ними возникает конкуренция. В таком случае вероятность успешного расселения падает.
Особенно наглядно это проявляется у хищников. Медведи и другие крупные хищники активно метят свои территории и агрессивно реагируют на любые попытки вторжения, особенно если конкурент относится к тому же или сходному по размерам виду. Мелкие хищники также метят территорию пахучими выделениями и вступают в схватки с другими хищниками своего размера. Например, лесная куница на границе своего ареала конкурирует с соболем, который обитает в том числе на севере Уральских гор.

Аналогичные примеры также встречаются среди птиц и грызунов. Восточный соловей на границе своего ареала конкурирует с западным соловьём: они занимают одну пищевую нишу и гнездятся похожим образом. Крапчатый и европейский суслики конкурируют в одной пищевой нише и сдерживают расселение вида-конкурента[22].

Экологические ограничения[править]
Территориальное распределение видов неоднородно. Оно существенно зависимости от особенностей среды и, главным образом, от доступности ресурсов, кртичиных для существования и развития популяции, которые распределены в большинстве случаев неравномерно. При этом пригодные для устойчивого развития того или иного вида территории могут находиться друг от друга на относительно небольшом расстоянии для миграции особей в течение их жизненного цикла (или переноса семян растений). Это даёт организмам возможность «испытывать» новые среды на предмет их пригодности для проживания, при наличии в них условий потенциально пригодных для жизни вида. Расселение является эффективным механизмом преодоления проблемы роста популяции и при наличии достаточного времени на адаптацию к изменениям условий окружающей среды, сопутствующим миграции, выступает в качестве фактора эволюционного развития вида . При наличии времени на адаптацию при расселения на новые территории у вида успевают вырабатываться фенотипические (например, поведенческие) или иногда генотипические изменения под новые, в ряде случаев экстремальные, условия проживания[23][24][25].
Например жук нарывник, Mylabris (Ammabris) elegantissima, зависит от песчаной акации, которая является его пищей. В итоге жук обитает в большинстве пустынь Средней Азии, но его нет в Ферганской долине, потому что там не растёт песчаная акация.[15]

Пальмовый гриф, хищная птица, относящаяся к семейству ястребиных. Однако взрослый пальмовый гриф на 60 % кормится растениями, особенно мякотью плодов масличной пальмы и рафии. Зависимость птиц от этих деревьев настолько высока, что их ареалы совпадают. Так же птицы питаются апельсинами, финиками, некоторыми злаками, семенами акации. Растительная пища у молодых птиц достигает 90 %. Когда особи взрослеют, они могут поедать крабов — морских и пресноводных, моллюсков, лягушек, головастиков, рыбу, навозных жуков, термитов, саранчу, мелких млекопитающих, птиц и их птенцов, змей, яйцами рептилий и даже падаль. Гнездование часто проводят на пальмах рафия, гнёзда строят у основания листьев.[26][27][28][29][30]
Одним из мощных побудительных факторов к миграции животных, как хищников, так и травоядных, является необратимое изменения климата. Резкое его изменение относительно скорости эволюционных механизмов создает проблемы для многих животных. Например, хохлатые пингвины к настоящему время расселены в нескольких климатических зонах (их географический ареал включает острова Субантарктики, Тасманию, Огненную Землю, материковое побережье Южной Америки). Этот вид имеет относительно большую продолжительность жизни — от 10 до 25 лет, что, с одной стороны, даёт возможность к миграции в зоны георгафического ареала с иными климатическими условиями, а с другой определяет низкие темпы микроэволюции вида за счёт медленной смены поколений. Выживание в условиях стремительно меняющегося климата за счёт быстрого приспособления при этом затруднено. В частности, у хохлатых пингвинов, живущих в различных климатических зонах велики поведенческие отличия. Проживание в субтропической зоне потребует от пингвинов субанатрктической зоны принципиально иных способов добывания пищи, а формирование таких поведенческих навыков займёт очень долгое время. Поэтому стратегией важивания хохлатых пингвинов при потепление в субарктической зоне будет не адаптация, а расселение в более пригодные для их проживания зоны Антарктики, что сопряжено с риском гибели многих особей при миграции[31][32].
Механизмы расселения животных и растений[править]
Механизмы расселения животных и растений определяются их образом жизни. Растения ведут неподвижный образ жизни, так как в процессе фотосинтеза способны использовать энергию солнечного света для превращения углекислого газа в питательные вещества, необходимые для роста и других жизненно важных процессов. Животные вынуждены получать пищу захватом питательных веществ напрямую из среды обитания. Неподвижные животные могут рассчитывать на получение пищи, приносимой потоками среды — главным образом морских и речных течений. Растения и неподвижные (малоподвижные) животные расселяются за счёт выброса в среду своих частей (пропагул), которые переносятся средой в новое место обитания. Животные, живущие на поверхности земли и в почве не погут обеспечить свою жизнедеятельность за счет пассивного захвата достаточного количества пищи. Способность к перемещению становится для них жизненно необходимым качеством, что делает в их случае поиск пищи и расселение связанными процессами.
Расселение растений[править]
Возможности растений по расширения своего ареала за счёт вегетативного размножения (частями побегов, корня и т. п.) ограничены. Растения расселяются за счет своих частей, называемых диаспорами, пропагулами растений или зачатками, которые отделяются естественным путём от материнского растения, далее распространяются под воздействием внешних условий, закрепляются на новых территориях с последующим возможным образованием новой популяции. К диспорам, ответственным за расселение, относятся семена, споры, плоды и соплодия, а также клубни и луковицы. Их распространение происходит за счёт абиотических и биотических факторов (векторов). Например, основные способы рассеивания семян включают абиотические — гравитация, ветер, вода и биотические векторы распространения — перенос животными, выброс (баллистический способ) семян из растения. Семена переносятся как самостоятельно, так и в составе плодов или вместе с остатками отмерших растений (перекати-поле). Кроме семян возможен перенос вегетативных диаспор в виде отдельных растений, например, ряска отпочковывает от себя дочерние растения, которые переносятся водоплавающими животными (рыбами, лягушками, птицами[33]). Существенный вклад в расселение растений также вносит хозяйственная деятельность человека[1].
Расселение животных[править]
- См. также: Океаническое расселение животных
Неподвижные животные[править]
Среди животных существет группа не способных к самостоятельному передвижению (немотильных) видов. Большинство из них ведут сидячий (прикреплённый) образ жизни — губки, мшанки, кораллы, устрицы. Некоторые из них (оболочники) ведут сидячий образ жизни во взрослой форме, но способны к перемещению в стадии личинки. В отдельных случаях (морские анемоны[34][35]) животное может открепляться от субстрата и медленно перемещаться (переползать) на новое место. Такие животные расселяются в основном пассивным способом: с течениями рек и морей, в том числе будучи прикреплёнными к плавающим предметам.
Интересным примером является расселение кораллов, которые при размножении согласованно (этот процесс согласуется с лунными циклами) выбрасывают сперматозоиды и яйцеклетки в воду. После их слияния получившаяся зигота быстро развивается в планулу, многоклеточное образование, которое сохраняет подвижность и затем оседает на субстрат для закрепления. Большая часть планул при этом погибает, оседая на неподходящие места или будучи съеденная зоопланктоном и донными хищниками (анемоны и другие кораллы). Но учитывая большое количество образовавшихся планул — десятки миллионов, часть их находит подходящий субстрат — голая известняковая порода и, оседая на ней, планула развивается до полипа. При дальнейшем развитии полип вырастает в коралловую головку, отпочковываются новые полипы, постепенно образуя новую колонию кораллов[36].
Подвижные животные[править]
Большинство видов животных являются мотильными, то есть способными к расселению за счёт собственных локомоторных способностей. Дальность расселения может быть различной для разных видов. Например, альбатросы способны к круглосуточным полётам и могут покрывать расстояния в тысячи километров, в то время как дальность однократного перелёта воробьев составляет 1-2 км. Наземные животные раселяются за счёт бега по земной поверхности и прыжков между растениям, белки-летяги могут перелетать между ветками и отдельными деревьями. Многие подвижные водоплавающие животные (рыбы, черепахи и др) при своих перемещениях активно используют течение воды. С плавающими стволами деревьев расселяются мелкие пресмыкающиеся — змеи, гекконы. Для мелких животных, таких как зоопланктон и отдельных водных насекомых, течение является доминирующим вектором расселения. Птицы и насекомые могут переносится при участии потоков ветра. Так, стая белых цапель была перенесена бурей из Африки в Америку, после чего заселила её. [1][37].
Расселение через стадию покоя[править]
Некоторых мелких животных расселяют более крупные, например, птицы, помимо паразитов, иногда переносят моллюсков, яйца пресноводных животных и др. Мелкие виды пресноводных беспозвоночных способны расселяться с помощью ветра или при переносе более крупными животными (птицами, млекопитающими или рыбами) в виде покоящихся яиц, эмбрионов или, в некоторых случаях, покоящихся взрослых стадий. Многие другие таксоны (кладоцеры, мшанки, гидры и др.) распространяются в виде покоящихся яиц или эмбрионов. Пресноводные губки обычно образуют специальные покоящиеся пропагулы — геммулы, служащие для расселения[1].
Многие из этих устойчивых покоящихся стадий животных — яйца, эмбрионы и взрослые особи — способны выдерживать не только засуху и экстремальные температуры (как низкие, так и высокие), но и воздействие пищеварительных ферментов при прохождении через желудочно-кишечный тракт птиц и других животных, высокую концентрацию солей и различные токсичные вещества. Наличие таких устойчивых форм обеспечивает возможность расселения на большие расстояния между водоёмами и способствует широкому распространению многих пресноводных и морских животных.
Расселение за счёт деятельности человека[править]
Иногда деятельность человека может расширять ареалы обитания, предоставляя новые способы распространения особей или семян. Примером является распространение животных с балластными водами судов. Ряд видов рыб и насекомых были перевезены таким образом на огромные расстояния и стали затем инвазивными видами в новых местах обитания[38].
Расселение животных в городах[править]
Воробьи живущие в Красноярске, в помещениях цехов и ТЭЦ, размножаются круглый год. Сроки размножения чёрной вороны, сизого голубя и большой синицы сдвинулись на 5-10 дней. Кряквы в городской черте Красноярска делают по 2-4 кладки за сезон, в природе кладка почти всегда одна[39].
Ранее дикие утки, жившие вне городов, активно адаптировались к постоянному проживанию в городах, даже в зимний период. Часть уток перестала улетать южнее, кормясь приносимыми людьми продуктами. Так же утки стали с одной стороны осторожнее, с другой не реагируют на фоновые шумы города — машины, работы техники, смех и крики людей. Они адаптировались к возможным угрозам от кошек и собак. Так, у крякв обитающий в Москве, дистанция вспугивания сократилась в 20 раз, по сравнению с утками обитающими в Московской области вне городов. Кряквы перестали совершать сезонны миграции, раньше приступают к гнездованию, размер выводка сократился[40][41].
Городские воробьи, изученные на Юго Западе Вирджинии, демонстрируют изменение поведения и способа размножения. Самцы этих воробьёв посещали свои гнёзда чаще, чем воробьи из сельских регионов, так же они раньше кормят своих птенцов. Так же в городах оказался выше гнездовой паразитизм, в частности буроголовый коровий трупиал занимает чужие гнёзда и откладывает свои яйца. Однако общий уровень хищничества в городах сильно ниже, чем в дикой местности и успех гнездования у птиц выше в пределах города[42][43].
В Москве воробьи ведут оседлый образ жизни, облигантно-синантропного типа, когда вид и паразитирует и соседствует с человеком. С 2000-х годов наблюдалось снижение их численности связанное с активным развитием парков, вместо диких насаждений. В период после 2020 года численность стабилизировалась, домовый воробей и полевой воробей адаптировались, кочуя зимой на переферию Москвы и возвращаясь в плотную застройку летом[44][45].
Последствия расселения[править]
[[Категория:Циклопедия:Незавершённый перевод с {{ЯЗЫК/{{{1}}}|р|на|=}}]]Файл:Lichen dispersal.ogv
Расселение имеет значение не только на уровне развития отдельных особей или отдельной популяции, но также может оказывать влияиние на экосистему в целом и вносить вклад в эволюционные процессы. Например, самая успешная в эволюционном смысле стратегия расселения для растений — перенос птицами за счет прикрепления семян или за счет их прогатывания. В итоге растения эффективно распространяются на десятки и порой сотни километров без собственных затрат энергии на перемещение. Благодаря такому расселению ареалы растений в течение истории Земли постоянно расширялись, насколько это позволяли географические и экологичестве факторы[46].
Для популяций животных, которые распределены по биому фрагментарно, расселение обеспечивает взаимосвязь субпопуляций в единую систему — метапопуляцию. Оно поддерживает обмен генами, что предохраняет отдельные популяции от инбридинга и снижает риск случайного вымирания в результате резкого изменения условий среды — если одна или несколько субпопуляций вымирают, велика вероятность повторного восстановления вида на данном участке местности за счёт быстрой колонизации.
Взаимная миграция особей между субпопуляциями снижает риск избыточной локальной адаптации, яркими примерами которой являются пути эволюционного развития коалы и большой панды, которые оказались на грани исчезновение. Считается, что панды около 2,6 млн лет назад утратили всеядность, присущую их предковому виду Ailurarctos lufengensis, который жил 7-8 млн лет назад, а около 5 тыс. лет рацион панд стал состоять только из отдельных видов бамбука[47][48][49].
Также как у панд, ареал коалы в последние несколько сотен тысяч лет существенно сузился и в настоящее время остался единственный вид рода каол. Причиной считается зависимость коал от нескольких видов эвкалипта, листья которых они исполозьзуют в пищу. Медлительность коал ведет к тому, что часто они полностью уничтожают побеги эвкалиптовых деревьев, на которых они проживают, что приводит к гибели растений. В итоге эвкалипт прутовидный также находится сейчас под угрозой исчезновения. Считается, что плотность популяции коал для их устойчивого развития должна составлять 1 особь на гектар эвкалиптового леса[50][51][52].
Расселение приводит к генетическим изменениям у многих видов. Так у отдельных видов пауков, обитающих на разных островах архипелага Канарских островов, наблюдалась взаимосвязь между расселением и накоплением генетических раличий внутри популяции — генетической дифференциации, которая в ряде случаев приводила к появлению новых подвидов — диверсификации[53].

Человек может оказывать сильное влияние на расселение животных, получаются опасные инвазивные виды. Например моллюски речная дрейссена, были эндемиками распространены в России. Ранее вид уже распространился, сначала по Дунаю и Днепру, затем попал в доки Великобритании, далее балластные воды кораблей захватывали личинки дрейссен и они попали в Великие озёра Северной Америки, став вредным инвазионным видом. На сегодня дрейссена расселилась до Калифорнии и попала в другие водоёмы США и Канады. Вред от этих моллюсков в том, что он активно размножается и засоряет водоочистные сооружения и водозаборы электростанций. Другой пример — завоз китайского пёстрого толстолобика и серебряного карпа, с целью борьбы с водорослями в прудах, в США. В итоге рыбы попали в реки Миссисипи, Миссури, Иллинойс и Огайо. Ряд видов вообще стал адаптироваться больше под проживание в городах и в целом среди людей, например воробьи, голуби, домовые мыши[54][55].
Были проведены исследования в Великобритании, на двух видах растений Капуста (род), как их семена переносятся ветром и людьми. Выяснилось что люди переносят семена намного дальше, чем мог перенести ветер. Семена, прикрепившиеся к обуви оставались на ней дор 8 часов и периодически отделялись, падая на землю[56].
Методы изучения расселения[править]
Для изучения расселения живых организмов применяются методы геногеографии, оценивающие влияние места проживания популяции на генетическое разнообразие. Они позволяют связать изменчивость популяции с климатом и особенностями ландшафта. Метод включает сбор образцов ДНК особей популяции и их сравнительный анализ. Для достаточно крупных животных отслеживание перемещения проводят с использованием датчиков GPS; в частности, такой метод активно применяется для изучения миграций птиц[57][58][59][60][61][62][63][64].
Иллюстрации[править]

Ареал тукана гвианская селенидера. Вид отделился от тукана tawny-tufted toucanet (Selenidera nattereri из-за реки.

Ареал обитания тукана tawny-tufted toucanet Selenidera nattereri.
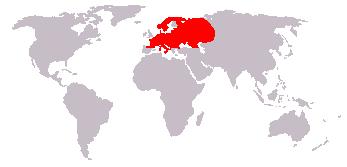
Ареал лесной куницы.

Ареал соболя.




Примечания[править]
- ↑ 1,0 1,1 1,2 1,3 1,4 1,5 1,6 БСЭ, 1975, Т. 21, с. 479.
- ↑ Расселение животных. www.booksite.ru. Проверено 28 апреля 2026.
- ↑ Меликян А.П., Девятов А.Г. Основные карпологические термины. — М.: Товарищество научных изданий КМК, 2001. — С. 19.
- ↑ Ophélie Ronce How Does It Feel to Be Like a Rolling Stone? Ten Questions About Dispersal Evolutionангл. // Annual Review of Ecology, Evolution, and Systematics. — 2007-12-01. — В. 1. — том 38. — С. 231–253. — ISSN 1543-592X. — DOI:10.1146/annurev.ecolsys.38.091206.095611
- ↑ Dries Bonte, Hans Van Dyck, James M. Bullock, Aurélie Coulon, Maria Delgado, Melanie Gibbs, Valerie Lehouck, Erik Matthysen, Karin Mustin, Marjo Saastamoinen, Nicolas Schtickzelle, Virginie M. Stevens, Sofie Vandewoestijne, Michel Baguette, Kamil Barton, Tim G. Benton, Audrey Chaput‐Bardy, Jean Clobert, Calvin Dytham, Thomas Hovestadt, Christoph M. Meier, Steve C. F. Palmer, Camille Turlure, Justin M. J. Travis Costs of dispersalангл. // Biological Reviews. — 2012-05. — В. 2. — том 87. — С. 290–312. — ISSN 1464-7931. — DOI:10.1111/j.1469-185X.2011.00201.x
- ↑ John B. Dunning, David J. Stewart, Brent J. Danielson, Barry R. Noon, Terry L. Root, Roland H. Lamberson, Ernest E. Stevens Spatially Explicit Population Models: Current Forms and Future Uses // Ecological Applications. — 1995-02. — В. 1. — том 5. — С. 3–11. — ISSN 1051-0761. — DOI:10.2307/1942045
- ↑ Metapopulation biology: ecology, genetics, and evolution / Ilkka Hanski, Michael E. Gilpin. — San Diego, CA: Academic Press, 2010. — ISBN 978-0-12-323446-9, 978-0-12-323445-2.
- ↑ Ilkka Hanski Metapopulation ecology. — Repr. — Oxford: Oxford University Press, 2005. — (Oxford series in ecology and evolution). — ISBN 978-0-19-854065-6.
- ↑ Emily K. Croteau, Edward J. Heist, Clayton K. Nielsen Fine-scale population structure and sex-biased dispersal in bobcats (Lynx rufus) from southern Illinoisангл. // Canadian Journal of Zoology. — 2010-06. — В. 6. — том 88. — С. 536–545. — ISSN 0008-4301. — DOI:10.1139/Z10-024
- ↑ Emily K. Croteau, Edward J. Heist, Clayton K. Nielsen Fine-scale population structure and sex-biased dispersal in bobcats (Lynx rufus) from southern Illinoisангл. // Canadian Journal of Zoology. — 2010-06. — В. 6. — том 88. — С. 536–545. — ISSN 0008-4301. — DOI:10.1139/Z10-024
- ↑ Emily K. Croteau, Edward J. Heist, Clayton K. Nielsen Fine-scale population structure and sex-biased dispersal in bobcats (Lynx rufus) from southern Illinoisангл. // Canadian Journal of Zoology. — 2010-06. — В. 6. — том 88. — С. 536–545. — ISSN 0008-4301. — DOI:10.1139/Z10-024
- ↑ Diana E. Bowler, Tim G. Benton Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamicsангл. // Biological Reviews. — 2005-05. — В. 2. — том 80. — С. 205–225. — ISSN 1464-7931. — DOI:10.1017/S1464793104006645
- ↑ Diana E. Bowler, Tim G. Benton Causes and consequences of animal dispersal strategies: relating individual behaviour to spatial dynamicsангл. // Biological Reviews. — 2005-05. — В. 2. — том 80. — С. 205–225. — ISSN 1464-7931. — DOI:10.1017/S1464793104006645
- ↑ Martin Kucera Jeder Fünfte macht zu viele unbezahlte Überstunden // kma - Klinik Management aktuell. — 2014-04. — В. 04. — том 19. — С. 17–17. — ISSN 1439-3514. — DOI:10.1055/s-0036-1577284
- ↑ 15,0 15,1 Гептнер В. Г. Общая зоогеография. — 1936.
- ↑ Hernani F. M. Oliveira, Marcella G. Santos, Renan J. Bosque, Larissa G. A. Goebel, Gabriela R. Longo, Daiana C. Silva, Priscilla L. Zangrandi, Guarino R. Colli, Fabricius M. C. B. Domingos How Do Rivers Divide Amazonian Animals? // Frontiers for Young Minds. — 2023-11-28. — том 11. — ISSN 2296-6846. — DOI:10.3389/frym.2023.1091144
- ↑ Naka, Luciano Nicolas Bird DNA helps explain Amazonian rivers’ role in evolutionen-US. The Conversation (2018-08-01). Проверено 29 апреля 2026.
- ↑ Moritz von der Lippe, Ingo Kowarik Do cities export biodiversity? Traffic as dispersal vector across urban–rural gradientsангл. // Diversity and Distributions. — 2008-01. — В. 1. — том 14. — С. 18–25. — ISSN 1366-9516. — DOI:10.1111/j.1472-4642.2007.00401.x
- ↑ Гептнер В. Г. Общая зоогеография. — 1936.
- ↑ Shalene Jha, C. Kremen Urban land use limits regional bumble bee gene flowангл. // Molecular Ecology. — 2013-05. — В. 9. — том 22. — С. 2483–2495. — ISSN 0962-1083. — DOI:10.1111/mec.12275
- ↑ Jonathan L. Richardson, Sozos Michaelides, Matthew Combs, Mihajla Djan, Lianne Bisch, Kerry Barrett, Georgianna Silveira, Justin Butler, Than Thar Aye, Jason Munshi‐South, Michael DiMatteo, Charles Brown, Thomas J. McGreevy Dispersal ability predicts spatial genetic structure in native mammals persisting across an urbanization gradientангл. // Evolutionary Applications. — 2021-01. — В. 1. — том 14. — С. 163–177. — ISSN 1752-4571. — DOI:10.1111/eva.13133
- ↑ Гептнер В. Г. Общая зоогеография. — 1936.
- ↑ Principles of conservation biology / Martha J. Groom, Gary K. Meffe, C. Ronald Carroll. — 3. ed. — Sunderland, Mass: Sinauer, 2006. — ISBN 978-0-87893-597-0, 978-0-87893-518-5.
- ↑ Thomas E. Martin [0656:AMPOCS2.0.CO;2 ARE MICROHABITAT PREFERENCES OF COEXISTING SPECIES UNDER SELECTION AND ADAPTIVE?]англ. // Ecology. — 1998-03. — В. 2. — том 79. — С. 656–670. — ISSN 0012-9658. — DOI:10.1890/0012-9658(1998)079[0656:AMPOCS]2.0.CO;2
- ↑ Monica Goigel Turner, R. H. Gardner, R. V. O'Neill Landscape ecology in theory and practice: pattern and process. — New York: Springer, 2001. — ISBN 978-0-387-95123-2, 978-0-387-95122-5.
- ↑ Palm-nut Vulture | The Peregrine Fund. peregrinefund.org. Проверено 29 апреля 2026.
- ↑ Anthony van Zyl Raptors of the world: a field guide by James Ferguson-Lees and David Christie // Ostrich. — 2006-11. — В. 3—4. — том 77. — С. 235–236. — ISSN 0030-6525. — DOI:10.2989/00306520609485540
- ↑ Peter Mikula, Federico Morelli, Radek K. Lučan, Darryl N. Jones, Piotr Tryjanowski Bats as prey of diurnal birds: a global perspectiveангл. // Mammal Review. — 2016-07. — В. 3. — том 46. — С. 160–174. — ISSN 0305-1838. — DOI:10.1111/mam.12060
- ↑ Camilo Carneiro, Mohamed Henriques, Castro Barbosa, Quintino Tchantchalam, Aissa Regalla, Ana R Patrício, Paulo Catry Ecology and behaviour of Palm-nut Vultures Gypohierax angolensis in the Bijagós Archipelago, Guinea-Bissauангл. // Ostrich. — 2017-08-31. — В. 2. — том 88. — С. 113–121. — ISSN 0030-6525. — DOI:10.2989/00306525.2017.1291540
- ↑ P. Holt MUNDY, P., BUTCHART, D., LEDGER, J. and PIPER, S. The vultures of Africa . Academic Press, London: 1992 (originally published by Acorn Books, Johannesburg: 1992). Pp 460; illustrated. Price: £55.00 ISBN: 0-12-510585-1англ. // Archives of Natural History. — 1994-02. — В. 1. — том 21. — С. 136–136. — ISSN 0260-9541. — DOI:10.3366/anh.1994.21.1.136
- ↑ Y Tremblay, Y Cherel Benthic and pelagic dives: a new foraging behaviour in rockhopper penguins // Marine Ecology Progress Series. — 2000. — том 204. — С. 257–267. — ISSN 0171-8630. — DOI:10.3354/meps204257
- ↑ Jaume Forcada, Philip N. Trathan Penguin responses to climate change in the Southern Oceanангл. // Global Change Biology. — 2009-07. — В. 7. — том 15. — С. 1618–1630. — ISSN 1354-1013. — DOI:10.1111/j.1365-2486.2009.01909.x
- ↑ Simona Paolacci, Marcel A. K. Jansen, Vlastimil Stejskal, Thomas C. Kelly, Neil E. Coughlan Metabolically active angiosperms survive passage through the digestive tract of a large-bodied waterbirdангл. // Royal Society Open Science. — 2023-03. — В. 3. — том 10. — ISSN 2054-5703. — DOI:10.1098/rsos.230090
- ↑ Актинии во льду • Картинка днярус.. «Элементы». Проверено 30 апреля 2026.
- ↑ (Actiniaria Hertwig, 1882) Отряд Актиниарии, Актинарии, Актинии, Морские анемоны, Морские цветы, Мясистые одиночные полипы, Order Actiniaria Hertwig, 1882 (Sea anemones, Actiniarias) 44 семейства. fishbiosystem.ru. Проверено 30 апреля 2026.
- ↑ Samuel C. Mogen, Nicole S. Lovenduski, Stephen G. Yeager, Antonietta Capotondi, Michael G. Jacox, Stephen Bograd, Emanuele Di Lorenzo, Elliot L. Hazen, Mercedes Pozo Buil, Who Kim, Nan Rosenbloom Multi-month forecasts of marine heatwaves and ocean acidification extremes // Nature Geoscience. — 2024-11-21. — В. 12. — том 17. — С. 1261–1267. — ISSN 1752-0894. — DOI:10.1038/s41561-024-01593-0
- ↑ Santiago Claramunt Flight efficiency explains differences in natal dispersal distances in birdsангл. // Ecology. — 2021-09. — В. 9. — том 102. — ISSN 0012-9658. — DOI:10.1002/ecy.3442
- ↑ Martin Kucera Beim Aufwand Spitze, beim Ergebnis Mittelfeld // kma - Klinik Management aktuell. — 2014-04. — В. 04. — том 19. — С. 17–17. — ISSN 1439-3514. — DOI:10.1055/s-0036-1577283
- ↑ «Утки в городе несутся трижды»: биолог о том, как меняется жизнь животных в Красноярском крае. newslab.ru. Проверено 28 апреля 2026.
- ↑ Почему городские утки не боятся никого — и как мегаполис сделал их умнее / Флора и фауна / iXBT Liveрус.. iXBT Live (2025-11-01). Проверено 28 апреля 2026.
- ↑ Совсем страх потеряли: московские утки отличаются от своих диких собратьеврус.. «Элементы». Проверено 28 апреля 2026.
- ↑ Сидельникова, Дарья Жизнь в городе делает самцов певчих воробьев «суперотцами»рус.. Naked Science (2023-08-22). Проверено 28 апреля 2026.
- ↑ Samuel J. Lane, Isaac J. VanDiest, Valerie N. Brewer, Courtney R. Linkous, Taylor E. Fossett, Christopher G. Goodchild, Kendra B. Sewall Indirect effects of urbanization: consequences of increased aggression in an urban male songbird for mates and offspringEnglish // Frontiers in Ecology and Evolution. — 2023-08-22. — том 11. — ISSN 2296-701X. — DOI:10.3389/fevo.2023.1234562
- ↑ Орнитологи МГУ объяснили, куда пропали воробьи в Москверус. (2025).
- ↑ Программа птицы Москвы и Подмосковья | Домовый воробей (Passer domesticus). birdsmoscow.net.ru. Проверено 28 апреля 2026.
- ↑ Geetha Ramaswami, Monica Kaushik, Soumya Prasad, Raman Sukumar, David Westcott Dispersal by generalist frugivores affects management of an invasive plant // Biotropica. — 2016-07-21. — В. 5. — том 48. — С. 638–644. — ISSN 0006-3606. — DOI:10.1111/btp.12343
- ↑ Добровидова, Ольга Панды перешли на бамбуковую диету не более 5 тысяч лет назадрус.. N + 1 — главное издание о науке, технике и технологиях. Проверено 28 апреля 2026.
- ↑ Han Han, Wei Wei, Yibo Hu, Yonggang Nie, Xueping Ji, Li Yan, Zejun Zhang, Xiaoxue Shi, Lifeng Zhu, Yunbing Luo, Weicai Chen, Fuwen Wei Diet Evolution and Habitat Contraction of Giant Pandas via Stable Isotope AnalysisEnglish // Current Biology. — 2019-02-18. — В. 4. — том 29. — С. 664–669.e2. — ISSN 0960-9822. — DOI:10.1016/j.cub.2018.12.051
- ↑ Ученые выяснили, когда панды перестали быть хищниками и стали есть бамбукрус.. Наука (2022-07-01). Проверено 28 апреля 2026.
- ↑ Русакова, Екатерина Генетики разобрались в любви коал к ядовитым листьям эвкалиптарус.. N + 1 — главное издание о науке, технике и технологиях. Проверено 28 апреля 2026.
- ↑ Ивтушок, Елизавета Контрацепция коал спасла эвкалипт от вымираниярус.. N + 1 — главное издание о науке, технике и технологиях. Проверено 28 апреля 2026.
- ↑ David S. L. Ramsey, Freyja Watters, David M. Forsyth, Matthew Wood, Charles R. Todd, Robyn Molsher, Phillip Cassey Long-term fertility control reduces overabundant koala populations and mitigates their impacts on food trees // Biological Conservation. — 2021-01-01. — том 253. — С. 108870. — ISSN 0006-3207. — DOI:10.1016/j.biocon.2020.108870
- ↑ Daniel Suárez, Paula Arribas, Eduardo Jiménez-García, Brent C. Emerson Dispersal ability and its consequences for population genetic differentiation and diversificationангл. // Proceedings of the Royal Society B: Biological Sciences. — 2022-05-25. — В. 1975. — том 289. — ISSN 0962-8452. — DOI:10.1098/rspb.2022.0489
- ↑ Emily K. Croteau, Edward J. Heist, Clayton K. Nielsen Fine-scale population structure and sex-biased dispersal in bobcats (Lynx rufus) from southern Illinoisангл. // Canadian Journal of Zoology. — 2010-06. — В. 6. — том 88. — С. 536–545. — ISSN 0008-4301. — DOI:10.1139/Z10-024
- ↑ Michael L. McKinney Urbanization as a major cause of biotic homogenizationангл. // Biological Conservation. — 2006-01. — В. 3. — том 127. — С. 247–260. — DOI:10.1016/j.biocon.2005.09.005
- ↑ Matthias C Wichmann, Matt J Alexander, Merel B Soons, Stephen Galsworthy, Laura Dunne, Robert Gould, Christina Fairfax, Marc Niggemann, Rosie S Hails, James M Bullock Human-mediated dispersal of seeds over long distancesангл. // Proceedings of the Royal Society B: Biological Sciences. — 2009-02-07. — В. 1656. — том 276. — С. 523–532. — ISSN 0962-8452. — DOI:10.1098/rspb.2008.1131
- ↑ Stéphanie Sherpa, Julien Renaud, Maya Guéguen, Gilles Besnard, Loic Mouyon, Delphine Rey, Laurence Després Landscape does matter: Disentangling founder effects from natural and human‐aided post‐introduction dispersal during an ongoing biological invasionангл. / Sharon Zytynska // Journal of Animal Ecology. — 2020-09. — В. 9. — том 89. — С. 2027–2042. — ISSN 0021-8790. — DOI:10.1111/1365-2656.13284
- ↑ Moritz von der Lippe, Ingo Kowarik Do cities export biodiversity? Traffic as dispersal vector across urban–rural gradientsангл. // Diversity and Distributions. — 2008-01. — В. 1. — том 14. — С. 18–25. — ISSN 1366-9516. — DOI:10.1111/j.1472-4642.2007.00401.x
- ↑ Luca Butikofer, Beatrix Jones, Roberto Sacchi, Marco Mangiacotti, Weihong Ji A new method for modelling biological invasions from early spread data accounting for anthropogenic dispersalангл. / Pedro Abellán // PLOS ONE. — 2018-11-27. — В. 11. — том 13. — С. e0205591. — ISSN 1932-6203. — DOI:10.1371/journal.pone.0205591
- ↑ Jean Clobert, Jean‐François Le Galliard, Julien Cote, Sandrine Meylan, Manuel Massot Informed dispersal, heterogeneity in animal dispersal syndromes and the dynamics of spatially structured populationsангл. // Ecology Letters. — 2009-03. — В. 3. — том 12. — С. 197–209. — ISSN 1461-023X. — DOI:10.1111/j.1461-0248.2008.01267.x
- ↑ Amin Vahedian Khezerlou, Xun Zhou, Xinyi Li, W. Nick Street, Yanhua Li DILSA+: Predicting Urban Dispersal Events through Deep Survival Analysis with Enhanced Urban Featuresангл. // ACM Transactions on Intelligent Systems and Technology. — 2021-08-31. — В. 4. — том 12. — С. 1–25. — ISSN 2157-6904. — DOI:10.1145/3469085
- ↑ Gabriele Cozzi, Nino Maag, Luca Börger, Tim H. Clutton‐Brock, Arpat Ozgul Socially informed dispersal in a territorial cooperative breederангл. / Garrett Street // Journal of Animal Ecology. — 2018-05. — В. 3. — том 87. — С. 838–849. — ISSN 0021-8790. — DOI:10.1111/1365-2656.12795
- ↑ Фахранурова, Лилия Ученые разобрались в навигации перелетных птицрус.. N + 1 — главное издание о науке, технике и технологиях. Проверено 28 апреля 2026.
- ↑ Л. В. Соколов [https://www.zin.ru/rybachy/sokolov-pdf/2011a.pdf СОВРЕМЕННАЯ ТЕЛЕМЕТРИЯ: НОВЫЕ ВОЗМОЖНОСТИ В ОРНИТОЛОГИИ]рус.. — 2011.
Литература[править]
- Большая советская энциклопедия : [рус.] : в 30 т. / под ред. А. М. Прохоров. — 3-е изд.. — М. : Советская энциклопедия, 1975. — Т. 21 : Проба – Ременсы. — 640 с.